
Business

Business
To improve the understanding of processes that control the distribution, occurrence, and behavior of gas hydrate systems over time, especially with respect to the role played by these systems in global climate change.
Idaho National Laboratory, Idaho Falls, ID 83415
Oregon State University, Corvallis, OR 97331
The current research is motivated by three hypotheses:
To test these hypotheses and to address parts of the current knowledge gap, four interrelated project objectives will be met. These include:
The results of this study will allow the scientific community to better place and understand the role of gas hydrates within the global carbon cycle. Gas hydrates are components of dynamic geobiological systems with inputs and outputs of methane, however, present rates of biological methane production are poorly understood. Efforts to model gas hydrate systems invariably ignore or parameterize these rates. The ultimate goal of this study is to produce significantly improved estimates for the rates of methanogenesis thereby increasing the accuracy of models pertaining to gas hydrates in the environment.
Work conducted under this project thus far has resulted in the development and implementation of sampling procedures that enable the evaluation of microbiological properties, the collection of microbiology samples from hydrate cores taken at onshore and offshore locations, the development of methodologies and tools for detecting and quantifying methanogens, and the publication of sample analyses and the insights these data have provided related to methanogenesis.
This integration of experimental and theoretical science represents a major step in the development of our understanding of gas hydrates in the environment. This project builds upon earlier work conducted by the principal investigators starting in 2002 (FWP-42C1-01). Major accomplishments of the earlier effort included:
The current research is divided into four interrelated tasks. Accomplishments to date, have focused on tasks 1, 2, and 4.
Task 1 – Methanogenesis rates at in situ conditions
The project is using biomass-recycling reactors (BRR) to cultivate Methanoculleus submarinus, a novel methanogen isolated from the Nankai Trough. Initial studies of M. submarinus determined that these cells reach a steady-state biomass after approximately 500 h (20 days). Between inoculation and day 20, cell biomass increased approximately 100-fold and then remained relatively constant for a year. Methanogenesis rate estimates show that the BRR is a useful tool for starving cells so that they can be used in experiments that seek to determine their activities when they are starved in their native environment.

M. submarinus has been sustained in the lab in a BRR for over one year. These cells are not growing and all of the substrate provided to the cells is directed towards cell maintenance. The starved cultures are used for experiments to determine the methane production rates (the amount of methane produced per cell during survival) under various seafloor conditions. The environmental conditions to which the M. submarinus culture are exposed include pressures (80-110 bars) and a range of dissolved product (i.e., methane) and substrate (i.e., energy-yielding compound like formate) concentrations. Initial estimates of methanogenic activity have been completed under starved conditions at 25oC. The affect of pressure on the cells using high-pressure vessels for the incubation of M. submarinus have been evaluated.
Studies of the tolerance of M. submarinus to pressure determined that the cells apparently exhibit enhanced catabolic activity when grown at pressures consistent with seafloor settings. Rates of methane formation were 2-4 times higher in cells incubated at 136 atm versus those incubated at 1 atm. In addition, carbon isotopic fractionation of the substrate was 6 per mil higher in the cells incubated at elevated pressures. The observed difference is likely from a kinetic effect related to the metabolic pathway used to make methane because the magnitude of equilibrium isotopic fractionation for abiotic chemical reactions Is not significantly affected by pressure. The increased kinetic isotopic fractionation may be due to restricted transport of substrates (e.g., formate, CO2) across the cell membranes at higher pressure or due to higher metabolic rates that the cells exhibit at higher pressure. Successive experiments at high pressure have been less conclusive and in fall 2008 we began to conduct basic physiological experiments aimed at determining fundamental metabolic capabilities of M. submarinus.
Task 2 – Molecular ecology of hydrate systems
In this task, methanogens and other microbial community members in sediments collected from hydrate-rich locations are being characterized and the key abiotic parameters that control biomass, diversity, and activity determined. Cores have been collected and analyzed from three separate field locations: 1) off the coast of India as a part of National Gas Hydrate Program (NGHP) research cruise on the R/V JOIDES Resolution during the Summer of 2006, 2) from beneath the permafrost at the Mt. Elbert Prospect well on the Alaskan North Slope during February 2007, and 3) from the Bullseye Vent and nearby active gas flares off the coast of Vancouver Island during the Pacific Geoscience Centre expedition (PGC2008007) in July-August 2008.
Microbial community diversity and their metabolic functioning with respect to primary carbon assimilation and respiration pathways from NGHP Site 17 were investigated. The samples were obtained from offshore India near the Andaman Islands in the Bay of Bengal, representing the deepest methane hydrate sediments found to date, likely due to the low geothermal gradient. The hydrate was found in sediment layers that contain coarse-grained volcanic ash in the depth range from 300 to 650 meters below the seafloor. DNA was extracted from 13 depth horizons, eight of which contained hydrates. Microscopic cell enumeration and domain-specific quantitative polymerase chain reaction (qPCR) revealed that those sediments harbor relatively low numbers of microbes that are composed mainly of Bacteria. Further functional gene amplification suggests that bacterial populations related to one-carbon (CO2 and CH4/CH3OH) biogeochemical cycling occur in the sediments examined. Compared to other seafloor settings, this Bacteria-dominated deep subseafloor habitat has unique phylogenetic and functional diversity.

Microbial communities collected from hydrate-bearing sediments on the ANS were studied to determine how abiotic variables (e.g., grain size, hydrate presence, original depositional environment) may control the type and distribution of microbes in the sediments. The cores were acquired from sub-permafrost, Eocene (35-36 million years ago [MYA]) sediments laid down as a marine transgressive series within which hydrates are believed to have formed 1.5 MYA. Forty samples, eight of which originally contained hydrates, were acquired from depths of ca. 606–666 meters below land surface. Five samples from drilling fluids acquired from the same depth range were included in the analysis as a control for contamination during the drilling and handling of cores. DNA was extracted from the samples and then amplified using polymerase chain reaction with primers specific for bacterial and archaeal 16S rDNA. Only bacterial DNA amplicons were detected. Further analysis revealed that sediment samples were distinct from those obtained from drilling fluids suggesting that the samples were not contaminated by the drilling fluids. All samples had evidence of microbial communities and sample depth, temperature, and hydrate presence appeared to have some influence on community diversity. Further examination of selected samples using clone libraries should help to identify the key taxa present in these unique sediments and yield a better understanding of the biogeochemistry of these gas-bearing systems.
Piston core samples were collected from Bullseye Vent with aim of obtaining the biofilm that was first observed in this location. This macroscopic pink to orange biofilm has been found offshore India and on the Cascadia Margin at Hydrate Ridge as well. Given that the biofilm appears to line sediment fractures where methane is present in abundance, it is thought that this is an as-yet unreported high concentration of cells sustained by this source of carbon and energy and that the biofilm may be involved in fundamental biogeochemical processes occurring within cold seep sites exhibiting variable methane and hydrocarbon fluxes. Biofilm and background sediment samples were preserved for later extraction of mRNA, DNA, and protein which can assist in determining microbial activities and relationships. Microbial analyses is expected to start in early FY2009.
Task 4 – Carbon cycle modeling
Collaboration with scientists at Rice University has yielded a one-dimensional numerical model that describes the formation and distribution of methane hydrate in marine sediment on geological time scales. The model examines the effect of changing seafloor temperature from 3 to 18 degree C in cases where microbial activity supplies most of the methane. Predictably, the temperature increase shifts the methane solubility curve in marine sediment and decreases the depth of the GHSZ. Less obvious but more important are temperature effects on the flux of seafloor organic carbon and the rate of methanogenesis. In some cases, increased seafloor temperature results in decreased amounts of methane hydrate. However, in other simulations, when seafloor organic fluxes and biogenic reaction rates increase significantly, amounts of methane hydrate can be higher than modeled for the present-day. It is possible that, during times of warm oceans, greater amounts of organic carbon enter the seafloor, microbes make methane from this carbon at much faster rates, and gas hydrate quantities exceed those at present-day. These somewhat counter-intuitive results may help to explain certain observations during warm climates.
Work continues on the following tasks:
Task 1 – Methanogenesis rates at in situ conditions
Cultures of Methanoculleus submarinus are being prepared for replication experiments to determine the effect of pressure on methanogen metabolic activity and carbon isotope fractionation. The intent is to conduct pressure experiments in an “accumulator” that can be used for repeated sampling (2.5- or 5.0-ml volumes) of pressure vessels without decompressing the cells in the system. Temperature incubation can be included with pressure as a variable. Incubation of the cells under high concentrations of methane will yield data on product inhibition and thermodynamic constraints used in Task 3. Experiments will also provide the pressure/methane production relationship that is required to refine the model that describes hydrate systematics in marine systems for Task 4. The results of these experiments will be integrated with the sample analyses from Task 2. By linking to Task 2 additional data will be gathered on the numbers of methanogens and other microbial communities and what controls their distribution in hydrate-bearing sediments. This will permit further understanding of methanogenic rates, the cycling of the molecule, and the impact of deep microbial communities on global carbon cycling and climate change. Task 2 will also explore the detection of methanogen-specific messenger RNA (mRNA) from cells that have been sustained in the BRR. Analysis of the mRNA will help to develop a molecular-based method for assessing methanogenic activity in sediments where methanogens demonstrate a range of metabolic activities.
Furthermore, experiments will be tightly connected to the thermodynamic modeling in Task 3 and that early iterations of this modeling effort will indicate the ratios of substrates and products that constrain methanogens to the limits of metabolic activity. Using this experimental approach, catabolic rates for this methanogen can be determined under conditions that permit initial extrapolation to hydrate environments from which we have determined the numbers of methanogens. The Task 4 carbon cycle modeling task will also be coordinated with this effort so that methanogenic metabolism an be tested under experimental conditions consistent with the range of conditions experienced by these microbes when they exist proximal to methane hydrate.
Currently, efforts to observe the metabolism of the Methanoculleus submarinus at elevated pressures continues to be difficult to perform because the cells are slow growing and prone to unexplained loss of viability. These studies at pressures consistent with seafloor settings have been difficult to conduct and are currently being re-evaluated for their feasibility to accomplish the intended objective.
Task 2 – Molecular ecology of hydrate systems
Work under this task will continue to characterize methanogens and other microbial community members in sediments collected from hydrate-rich locations and determine the key abiotic parameters that control biomass, diversity, and activity. Sites that will be focused on include: 1) offshore India (cored: 2006); 2) Mt. Elbert well on the Alaskan North Slope (cored: 2007); and 3) Bullseye Vent on the Cascadia Margin (cored: 2008).
For each of these sites, the physical and chemical factors that control microbial community distribution and methanogen biomass will be assessed. Multivariate statistical analyses of the microbial community signatures that occur in the sediments will be performed with respect to various abiotic properties of the sediments including presence/absence of hydrates, grain size of the sedimentary materials, original depositional environment. Results will be compared to other findings from subsurface strata, both terrestrial and marine and then integrated with the computational models that describe methanogenesis in sub-seafloor and sub-permafrost sediments (Tasks 3 and 4). The results obtained will also be used to inform future sampling efforts or microbial studies in strata where hydrates are present.
Recent efforts have focused on developing clone libraries representative of the microbial communities from some of the Mt Elbert samples. Characterization of the microbial communities using terminal-restriction fragment length polymorphism (t-RFLP) combined with non-metric multidimensional scaling (NMS) has allowed a determination of abiotic variables that influence bacterial diversity in these sediments. The key abiotic variables that appear to structure the microbial communities in the Mt. Elbert sediments are shown in the figure below.
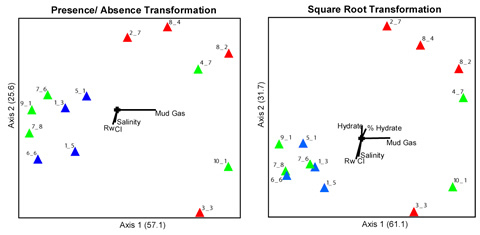
2-dimensional NMS ordination of sample units from the Mt Elbert well for presence/absence (P/A) and square root transformations. The blue triangles are samples that lacked hydrate, the red triangles contained hydrate, and the green triangles are samples that were found at boundaries between hydrate and non-hydrate zones. The amount of variance explained by each axis appears in parentheses with the total variance explained by the first two axes for the P/A transformation and square root transformation being 82.7% and 92.8%, respectively. The vectors show the direction and magnitude of correlation between sample units and selected environmental parameters measured in the samples. Rw = resistivity of the sediment, Cl = chloride concentration, Salinity = salt concentration, Mud Gas = concentration of methane gas in the drilling fluids, % Hydrate = the percentage of pore space that is occupied by hydrate, Hydrate = presence or absence of hydrate in the sample.
Investigation into the the molecular ecology of a pigmented biofilm found in relatively shallow (<10 mbsf) sediments in hydrate coring efforts around the world is also continuing. This macroscopic pink to orange biofilm has been found offshore India and on the Cascadia Margin at both Hydrate Ridge and off Vancouver Island. The biofilm occurs in sediment fractures in seafloor settings where methane is present in abundance and it is suspected that the biofilm communities depend on the methane as a source of carbon and energy. Biofilm and background sediment samples were collected as a part of a Pacific Geoscience Center cruise to Barclay Canyon during July-August 2008. Samples were preserved with RNALater that will permit the extraction of mRNA, DNA, and protein allowing us to determine the microbial activities and relationships present in the biofilms as well as the surrounding sediments.
Task 3 - Thermodynamic and kinetic modeling
This task awaits further progress from Task 1. W. Ussler at Monterey Bay Aquarium Research Institute (MBARI) has developed preliminary thermodynamic models that address the biogeochemical reactions associated with microbial methane production and oxidation in deep-sea sediments. These models will be tailored to the specific laboratory conditions used in Task 1, with attention towards determining the theoretical threshold at which methanogenesis can occur and simulating the effects pressure and substrate availability have on microbial metabolism. As part of this collaborative effort, realistic kinetic models for high pressure microbial metabolism will be developed and coupled with the thermodynamic models to produce a better framework for designing and evaluating the laboratory incubations performed under Task 1 and for application of these results to gas hydrate formation and stability, and carbon cycling in the geologic environment on geologic time-scales (Task 4). To the extent possible, findings from the molecular ecology work (Task 2) will be integrated into considerations of where in sediments such biogeochemical reactions are likely or unlikely to occur.
Work under this task has also contributed to the recent release of the model from the Mineral Management Service entitled: Preliminary Evaluation of In-Place Gas Hydrate Resources: Gulf of Mexico Outer Continental Shelf (MMS-2008-004) [PDF -external site]. In the report, constraints were considered that would be experienced by methane producing archaea under conditions of high methane concentration and how their activity may be controlled by the methane. The model also includes consideration of how temperature controls biological production of methane in Gulf of Mexico sediments and where methanogenesis is likely to be limited by high temperatures.
Also pertinent to this task, the PI has co-authored a recent manuscript (Caldwell et al. 2008. Environ. Sci. Technol. 42: 6791-6799) that considers thermodynamic conditions in marine sediments where methane is generated and consumed, and the PI and Ussler have completed a chapter (Colwell, F.S. and W. Ussler. In: Handbook of Hydrocarbon Microbiology. Springer-Verlag, Inc. In press.) that considers the detrimental consequences of of hydrocarbon:microbe interactions from a global perspective. This chapter keys largely on the biological production of methane in seafloor and permafrost settings.
Task 4 - Carbon cycle modeling
In this task the results from the previous tasks will be used to drive a one-dimensional numerical model developed at Rice University that describes the formation and distribution of methane hydrate in marine sediment systems. The model considers how variable seafloor temperature would alter the methane solubility curve leading to a decrease in the depth of the hydrate stability zone, and temperature effects on the flux of seafloor organic carbon and the rate of methanogenesis. A manuscript, “Elevated bottom water temperature and abundant marine gas hydrate in the early Palaeogene", was submitted to Nature-Geoscience in October 2009.
This field work proposal was discontinued in December 2009 and work by the PI, Rick Colwell, is now integrated into NETL in-house research efforts.
$940,000
$0
NETL – Robert Vagnetti (robert.vagnetti@netl.doe.gov or 304-285-1334)
Oregon State University – Rick Colwell (rcolwell@coas.oregonstate.edu or 541-737-5220)
In addition to the information provided here, a full listing of project related publications and presentations as well as a listing of funded students can be found in the Methane Hydrate Program Bibliography [PDF].
2008 ICGH Paper - Distribution of the Dominant Microbial Communities in Marine Sediments Containing High Concentrations of Gas Hydrates [PDF]
Fire in the Ice article, [PDF-883KB] " Methanogenesis in hydrate-bearing sediments: integration of experimental and theoretical approaches" by Colwell, F., W. Ussler, and G. Dickens, 2006 - Winter edition - 2006, pg. 1
Pertinent Publications
F. Colwell, A. Hangsterfer, E. Brodie, R. Daly, M. Holland, B. Briggs, P. Carini, M. Torres, M. Kastner, P. Long, H. Schaef, M. Delwiche, W. Winters, “Fine scale control of microbial communities in deep marine sediments that contain hydrates and high concentrations of methane,” [PPT - 5.19MB] Moscone Center, San Francisco, CA; American Geophyscial Union, Fall Meeting; Dec 10-14, 2007.
Colwell, F., T. Nunoura, M. Delwiche, S. Boyd, R. Bolton, D. Reed, K. Takai, R. Lehman, K. Horikoshi, D. Elias, and T. Phelps, 2005, Evidence of minimal methanogenic numbers and activity in sediments collected from the JAPEX/JNOC/GSC et al. Mallik 5L-38 gas hydrate production research well: in Dallimore, S. and T. Collett, eds., Scientific Results from the Mallik 2002 Gas Hydrate Production Research Well Program, Mackenzie Delta, Northwest Territories, Canada, Geological Survey of Canada Bulletin, Bulletin 585, p. 102.
Colwell, F., and R. Smith, 2004, Unifying principles of the deep terrestrial and deep marine biospheres, in Wilcock, W., E. Delong, D. Kelley, J. Baross, and S. Cary, eds., Subseafloor Biosphere at Mid-Ocean Ridges: Geophysical Monograph Series, v. 104. American Geophysical Union, Washington, D.C., p. 355-367.
Colwell, F., M. Delwiche, D. Blackwelder, R. Cherry, J. Mikucki, Y. Liu, D. Boone, and T. Uchida, 2002, Evidence of broad thermal tolerance of methanogens in sediments containing gas hydrates. Proceedings of the 4th International Conference on Gas Hydrates, Yokohama, Japan, p. 19-24.
Colwell, F., M. Delwiche, D. Blackwelder, M. Wilson, R. Lehman, and T. Uchida, 1999, Microbial communities from core intervals, JAPEX/JNOC/GSC Mallik 2L-38 gas hydrate research well: in Dallimore, S., T. Uchida, and T. Collett, eds., Scientific Results from JAPEX/JNOC/GSC Mallik 2L-38 Gas Hydrate Research Well, Mackenzie Delta, Northwest Territories, Canada: Geological Survey of Canada, Bulletin 544. pp. 189-195
Colwell, F., R. Matsumoto, and D. Reed, 2004, A review of the gas hydrates, geology and biology of the Nankai Trough. Chemical Geology, v. 205, p. 391-404.
Inagaki, F., T. Nunoura, S. Nakagawa, A Teske, M. Lever, A. Lauer, M. Suzuki, K. Takai, M. Delwiche, F. Colwell, K. Nealson, K. Horikoshi, S. D’Hondt, and B. Jørgensen, 2006, Biogeographical distribution and diversity of microbes in methane hydrate-bearing deep marine sediments on the Pacific Ocean Margin. Proc. Nat. Acad. Sci. USA. 103: 2815-2820.
Lorenson, T., F. Colwell, M. Delwiche, and J. Dougherty, in review, Acetate and hydrogen concentrations in pore fluids associated with a large gas hydrate reservoir, offshore Oregon, USA. Ocean Drilling Program, Leg 204, Data Reports.
Lorenson, T.D., F.S. Colwell, M. Delwiche, and J.A. Dougherty. 2006. Data report: Acetate and hydrogen concentrations in pore fluids associated with a large gas hydrate reservoir, southern Hydrate Ridge, offshore Oregon, USA. In Tréhu, A.M., Bohrmann, G., Torres, M.E., and Colwell, F.S. (Eds.), Proc. ODP, Sci. Results, 204 [Online]. Available from World Wide Web: http://www-odp.tamu.edu/publications/204_SR/126/126.htm [external site].
Mikucki, J., Y. Liu, M. Delwiche, F. Colwell, and D. Boone, 2003, Isolation of a Methanogen from deep marine sediments that contain methane hydrates, and description of Methanoculleus submarinus sp. nov. Applied and Environmental Microbiology, v. 69, p. 3311-3316.
Reed, D., Y. Fujita, M. Delwiche, D. Blackwelder, P. Sheridan, T. Uchida, and F. Colwell, 2002, Microbial communities from methane hydrate-bearing deep marine sediments in a forearc basin. Applied and Environmental Microbiology, v. 68, p. 3759-3770.
Trehu, A.M., M.E. Torres, G. Bohrmann, and F.S. Colwell. 2006. Leg 204 synthesis: Gas hydrate distribution and dynamics in the central Cascadia accretionary complex, pgs 1-40. In (A.M. Trehu, M.E. Torres, G. Bohrmann, and F.S. Colwell, eds.). Proc. ODP, Sci. Results, 204: College Station, TX (Ocean Drilling Program), doi:10.2973/odp.proc.sr.204.101.2006; http://www-odp.tamu.edu/publications/204_SR/synth/synth.htm [external site]
Presentations
Boyd, S., et al., 2004, Determination of methanogen biomass in hydrate-bearing sediments by quantitative PCR: Annual Meeting of the American Society for Microbiology Annual Meeting, New Orleans, LA. May.
Colwell, F., S. Boyd, M. Delwiche, and D. Reed, 2005, Rates of biological methane production in sediments: 4th International Workshop on Gas Hydrates, Victoria, BC.
Colwell, F., T. Lorenson, S. Boyd, M. Delwiche, D. Reed, and D. Newby, 2005, Experimentally determined rates of methanogenesis in methane-bearing sediments: Geological Society of America Annual Meeting, Salt Lake, UT. October 16-19.
Colwell, F., S. Boyd, M. Delwiche, and D. Reed, 2004, Realistic Rates of Biological Methane Production in Hydrate Bearing Sediments: The American Association of Petroleum Geologists Hedberg Conference, Vancouver, BC., September 12-16.
Colwell, F., M. Delwiche, D. Reed, S. Boyd, T. Nunoura, K. Takai, and the ODP Leg 204 Shipboard Scientific Party, 2003, Microbiology of hydrates: searching for the methane source term: Goldschmidt Conference, Kurashiki, Japan, September.
Colwell, F., et al, 2003, Microorganisms of methane hydrates: Down but not out: Annual Meeting of the American Association for the Advancement of Science (AAAS), Denver, CO. February.
Colwell, F., M. Delwiche, D. Reed, S. Boyd, T. Nunoura, F. Inagaki, and K. Takai, 2003, Geomicrobiology and methanogenesis in accretionary complexes: American Geophysical Union Fall Meeting, San Francisco, CA. December.
Colwell, F., et al., 2003, Microorganisms of methane hydrates: Down but not out: Goldschmidt Conference, Kurashiki, Japan. September. Keynote address.
Colwell, F., D. Reed, Y. Fujita, M. Delwiche, D. Blackwelder, T. Uchida, T. Fujii, and H. Lu, 2001, Deep-subsurface marine methane hydrate microbial communities: Who’s there and what are they doing?: American Geophysical Union, Fall Meeting, San Francisco, CA. December.
Colwell, F., M. Dewiche, D. Blackwelder, Y. Fujita, and T. Uchida, 2000, Microbial methane production and biomass in the Nankai Trough: Western Pacific Geophysics Meeting, Tokyo, Japan. June.
Colwell, F., 1999, Subsurface microbial communities in sediments bearing methane hydrates: Engineering Foundation Meeting, Salt Lake City, UT. July.
Inagaki, F., T. Nunoura, M. Suzuki, K. Takai, K. Nealson, K. Horikoshi, M. Delwiche, F. Colwell, and B. Jorgensen, 2003, Microbial community structures in methane hydrate bearing deep marine sediments form the Peru Margin (ODP Leg 201) and the Cascadia Margin (ODP Leg 204): American Geophysical Union, Fall Meeting, San Francisco, CA, December.
Nunoura, T., K. Takai, K. Nealson, K. Horikoshi, M. Delwiche, F. Colwell, and ODP leg 204 Shipboard Scientific Party, 2003, The microbial community structure of methane hydrate bearing deep subseafloor sediments in Cascadia Margin (ODP Leg204): Goldschmidt Conference, Kurashiki, Japan. September.

